引言
有效的过继性T细胞疗法(ACT)包括通过治疗性使用转移的T细胞来杀伤癌细胞。主要的ACT方法之一是嵌合抗原受体(CAR)T细胞疗法。CAR T细胞通过使T细胞能够通过单链可变片段(scFv)识别域结合靶细胞表面抗原,介导MHC非限制性的肿瘤细胞杀伤。结合后,CAR T细胞形成非经典的免疫突触(IS),这是其效应功能所必需的。这些细胞随后通过穿孔素和颗粒酶轴、Fas和Fas配体轴,以及释放细胞因子以敏化肿瘤基质,来介导其抗肿瘤效应。它们在宿主体内的持久性和功能输出严格依赖于受体各个组件——scFv、间隔区结构域和共刺激结构域——以及这些组件的功能如何协同作用以增强CAR T细胞的性能。
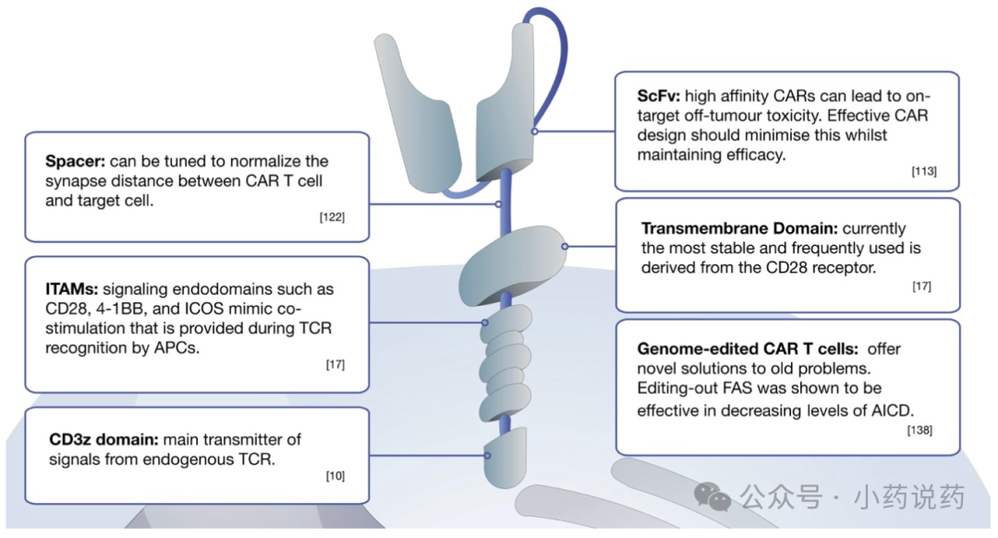
CAR是一种合成结构,可以通过单链可变片段(scFv)识别域结合靶细胞表面抗原。最初的概念将这个配体识别域与一个由CD3ζ链的一部分组成的细胞内信号模块连接起来,以在抗原结合后诱导T细胞激活。这两个模块通过一个细胞外铰链结构域和一个跨膜结构域连接,形成了最简单的CAR形式,被称为第一代CAR。TCR CD3ζ链包含3个免疫酪氨酸激活基序(ITAMs),因此,即使没有TCR-CD3复合物的其他组分(γ、δ和ε链),该链本身也能传递强大的信号。信号由淋巴细胞特异性蛋白酪氨酸激酶(Lck)介导的CD3胞质域内ITAMs的磷酸化启动。
后续改进现有CAR分子的努力导致了第二代和第三代CAR结构的出现,这些结构整合了信号内结构域,如CD28、CD137(也称为4-1BB)和诱导性T细胞共刺激因子(ICOS),试图模拟抗原呈递细胞(APCs)在TCR识别期间提供的共刺激。这种由磷酸肌醇3-激酶PI3K(在CD28的情况下)传播的共刺激信号,对于完全的生理性T细胞激活是必需的。向第四代或第五代CAR T细胞的进一步发展包括了来自细胞因子受体的信号结构域或炎症细胞因子(如IL-12或IL-18)的可诱导表达。
-02-
一、非经典免疫突触形成
传统的细胞毒性T淋巴细胞(CTLs)由于其TCR结合后形成独特的免疫学突触,能够迅速破坏并消除其靶细胞。这种高度有组织的结构由一系列同心环,即超分子激活簇(SMAC)组成,每个环源自赋予特定功能的聚集分子。中央SMAC(cSMAC)由TCR和Lck簇组成,通过积累T细胞激活信号和递送细胞毒性颗粒来增强和放大致死反应。它被外周SMAC(pSMAC)包围,pSMAC是淋巴细胞功能相关抗原-1(LFA-1)粘附分子的一个环,可稳定IS和靶细胞结合。远端SMAC(dSMAC)是肌动蛋白的聚集,完成了IS的靶心结构。
已知CAR T细胞至少部分利用常规的TCR信号传导机制。因此,人们可能认为CAR T细胞-靶细胞IS与经典的相似。然而,研究揭示了CAR形成的IS结构与经典TCR IS相比存在显著差异。虽然证明了LFA-1对于两种受体的IS形成都是可有可无的,但CAR IS缺乏cSMAC内Lck的聚集,并显示出更无序的Lck微斑模式。因此,发现CAR IS的内径尺寸显著更小,这与更快的CAR T细胞脱离相关。此外,大规模反相蛋白阵列鉴定出CAR T细胞中近端信号蛋白蛋白激酶C-delta(PKCδ)的快速下调,表明CAR启动的信号持续时间更短。这些发现得到了向IS加速递送裂解颗粒的支持,导致更快的靶细胞杀伤。
形成稳定的IS是T细胞诱导靶细胞杀伤的主要前提。一旦IS形成,效应细胞可以利用不同的途径诱导肿瘤细胞裂解。
-03-
二、穿孔素和颗粒酶
为了介导细胞溶解效应功能,T细胞主要利用两种主要途径:含有穿孔素和颗粒酶的细胞毒性颗粒的胞吐作用,以及膜结合肿瘤坏死因子(TNF)家族配体的表达,这些配体与其各自的受体结合后诱导靶细胞凋亡。这两种途径可分为慢作用(TNF家族配体)和快作用(脱颗粒)杀伤机制。
为了确保快速而精确地杀伤感染或恶性靶细胞,细胞毒性颗粒锚定在效应细胞的微管上。在免疫突触形成后,颗粒向界面迁移并在cSMAC区域内与质膜融合。含有其细胞溶解有效载荷的囊泡被释放到突触间隙中,在那里穿孔素诱导靶细胞膜上的孔形成,以促进促凋亡颗粒酶的进入。一旦进入靶细胞的细胞质,颗粒酶可以通过切割其关键底物来诱导caspase依赖性和非依赖性的凋亡性细胞死亡。
然而,在小鼠系统中,CD8⁺细胞毒性T淋巴细胞依赖于颗粒胞吐作用,而CD4⁺ CTLs则利用Fas和Fas配体(FasL)途径来介导细胞毒性,而在人类系统中,CTLs(包括CD8⁺和CD4⁺)主要利用细胞溶解的穿孔素和颗粒酶轴来介导靶细胞凋亡。此外,当配备CAR时,两个亚群的人类T细胞都可以以MHC和Fas独立的方式有效根除肿瘤细胞。因此,穿孔素和颗粒酶的细胞溶解脱颗粒被认为是CAR T细胞发挥重定向靶细胞杀伤的主要机制。
此外,CD4⁺ T细胞与CD8⁺ T细胞相比,细胞内穿孔素和颗粒酶的表达量显著较低。因此,CD4⁺ CAR T细胞的有效靶细胞杀伤要么被延迟,要么需要更多的效应细胞数量才能达到与CD8⁺ CAR T细胞相当的细胞溶解。
-04-
三、Fas和Fas配体(FasL)轴
Fas和FasL途径由Fas受体被Fas配体三聚化引导。这导致caspase 8的激活(由衔接蛋白Fas相关死亡结构域(FADD)和pro-caspase 8介导,它们形成死亡诱导信号复合物(DISC))。活化的caspase 8然后负责下游pro-caspase 3的加工,形成成熟的caspase 3,caspase 3通过随后切割超过500个细胞底物继续介导细胞死亡,有效地执行凋亡程序。研究表明,活化的CD30和CD19靶向CAR T细胞能够以抗原独立、细胞-细胞接触介导的方式介导针对抗原阴性部分的肿瘤裂解。
总之,Fas和FasL轴构成了CAR T细胞在异质性肿瘤环境中介导肿瘤细胞裂解的另一种机制途径。抗原非依赖性的旁观者杀伤机制提供了一条可以在抗原丢失相关疾病复发背景下利用的途径。此外,由于治疗相关毒性的挑战仍未解决,利用这一途径可能有助于克服与穿孔素和颗粒酶介导的细胞毒性失败相关的并发症。
四、细胞因子产生
已经表明,HER-2特异性CAR T细胞衍生的细胞因子可以诱导肿瘤基质上的干扰素γ受体表达,同时驱动免疫细胞再教育(例如将巨噬细胞极化为抗肿瘤M1表型)。除了抗原特异性肿瘤细胞靶向外,CAR T细胞驱动的抗原非依赖性基质破坏突出了CAR T细胞的另一种作用模式。
另外一个系统是使用用于通用细胞因子杀伤的重定向T细胞(TRUCKs)。这些CAR T细胞被用作在肿瘤组织内分泌和介导效应细胞因子积累的载体。这种方法允许在肿瘤组织内控制和定点递送效应分子,避免了其全身性递送所遇到的问题。例如一种具有多效性功能的分子IL-12,可以激发抗肿瘤免疫反应。机制包括改善T细胞细胞溶解活性,募集和激活先天免疫细胞,以及重编程基质相关的免疫抑制细胞。TRUCK方法依赖于通过CAR重定向T细胞递送IL-12,同时以MHC非限制性方式靶向肿瘤相关抗原,实现了这一点。
五、CAR T细胞作为连续杀伤者
理想情况下,ACT将以相对较少的效应细胞数量介导肿瘤块的快速破坏,确保更好地控制副作用。自然细胞毒性淋巴细胞(如NK和CD8⁺ T细胞)连续杀伤多个靶细胞的能力已被证明。同样,CAR T细胞的连续杀伤潜力也得到了验证。
另一个重要方面是CAR T细胞产品中CD8⁺和CD4⁺ T细胞的比例。缺乏CD4⁺ T辅助细胞可导致CD8⁺ T细胞功能障碍和凋亡。通过将CD4⁺ T细胞纳入治疗方案,它们在提供有效免疫疗法中的重要性已经变得明显。这两种细胞类型之间在动力学和机制水平上的相互作用的重要性,通过观察抗CD19 CAR CD8⁺和CD4⁺ CAR T细胞之间的纵向相互作用进行了研究。单细胞分析显示,CD4⁺和CD8⁺ CAR T细胞具有同等的肿瘤细胞杀伤能力,尽管前者需要更长的结合时间和延迟的动力学。
总的来说,很明显,CD4⁺ CAR T细胞介导杀伤的潜力可以进一步增强CAR T细胞疗法的疗效。展望未来,需要确定和提供最佳的CAR T细胞亚群组成,这一领域已经成为当前临床研究的焦点。
六、CAR设计的亲和力变化可以最大化杀伤效率
许多CAR T细胞方法使用具有高亲和力的scFvs。这主要基于先前的研究,这些研究表明CAR T细胞激活阈值与scFv亲和力呈负相关。研究TCR刺激发现,存在一个理想的T细胞激活亲和力窗口,因为进一步的TCR亲和力增强并未改善治疗效果。与抗体或抗体偶联药物相比,CAR T细胞方法具有更高的靶标灵敏度极限(几个数量级)。由于在肿瘤部位特异性表达的肿瘤靶点如此之少,亲和力调节需要更有效地用于更安全、更可控的CAR T细胞疗法。
虽然scFv亲和力对于CAR功能和增强杀伤潜力显然很重要,但为了优化功能,还必须考虑CAR设计的其他组成部分。
七、优化CAR T细胞功能和杀伤潜力
为CAR T细胞介导的杀伤接合替代信号通路在临床前研究中证明非常有效。例如一种旨在激活Janus激酶-信号转导和转录激活因子(JAK-STAT)通路的CAR构建体。他们可以通过整合截短的IL-2受体β(IL2RB)链来激活STAT5,并通过在CD3z的C末端添加一个YXXQ基序来激活STAT3。这种CAR触发了类似IL-21处理的基因表达谱,使T细胞具有独特的功能,包括改善的增殖能力和优于CD28-CD3z和4-1BB-CD3z CAR T细胞的效应功能。
这些差异得到了进一步探索。在弥漫性淋巴瘤异种移植模型中,CD28-CD3z CAR显示出CAR CD3z链和CAR相关Lck的基础磷酸化增加,导致更强的动力学和信号强度,这与效应T细胞样表型和功能相关。这种信号强度部分归因于Lck与该结构域在CAR复合物中的组成性关联。相比之下,4-1BB-CD3z CAR T细胞优先表达T细胞记忆相关基因,并在体内对已形成的肿瘤表现出持续的抗肿瘤活性。
对CAR分子本身进行的优化是该领域一个持续的学习过程。随着新的识别域或共刺激域星3式的出现,可能需要重新审视先前的优化参数。
结语
CAR-T细胞疗法代表了癌症免疫治疗领域的一项革命性突破,尤其在血液系统恶性肿瘤中已展现出卓越的临床疗效。其核心作用机制依赖于CAR T细胞与靶细胞形成非经典免疫突触后,通过穿孔素/颗粒酶轴、Fas/FasL轴以及细胞因子分泌等多重途径介导高效、精准的肿瘤细胞杀伤。然而,该疗法在实体瘤中的应用仍面临巨大挑战,包括肿瘤微环境的抑制、靶抗原异质性以及治疗相关毒性等。
尽管挑战犹存,但CAR T细胞疗法模块化、可编程的特性为其持续进化提供了无限可能。随着对CAR T细胞生物学机制的更深入理解,以及人工智能辅助设计等前沿技术的融合,下一代CAR T疗法有望在提升实体瘤疗效、管理毒性及实现个性化治疗方面取得根本性进展,最终为更广泛的癌症患者带来治愈希望。
参考资料:
Killing Mechanisms of Chimeric Antigen Receptor (CAR) T Cells. Int J Mol Sci. 2019 Mar 14;20(6):1283.

以上直播课和PPT均可在知识星球中免费下载,扫描下方二维码加入“小药说药知识文库”的知识星球

公众号已建立“小药说药专业交流群”微信行业交流群以及读者交流群,扫描下方小编二维码加入,入行业群请主动告知姓名、工作单位和职务。
