引言
今天我们要重点聊聊适应性免疫系统里最重要的成员之一:B细胞。像所有血细胞一样,B 细胞来源于骨髓,从干细胞中分化而来。在人类的整个生命周期中,每天大约能够产生10亿个B细胞,即使是老年人也能源源不断产生新的B细胞。
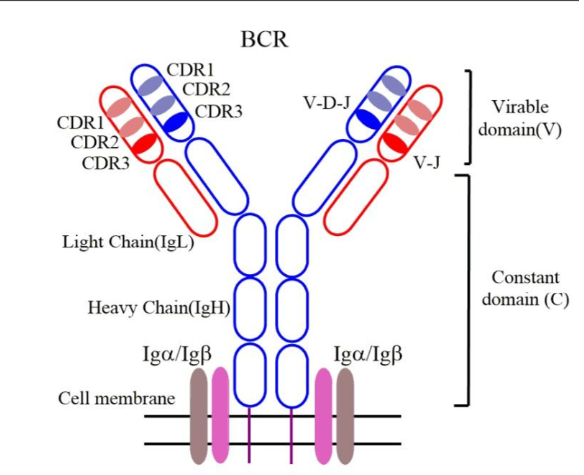
在B细胞的表面有一个重要的结构,B细胞受体(BCR),它们的结构与抗体分子几乎相同,唯一的区别是抗体的重链末端缺乏锚定于细胞膜的蛋白序列。由于缺乏这种序列,抗体分子不能够锚定在B细胞表面,而会被运出B细胞,从而自由地到达全身各处行使职责。
B细胞受体:组合的艺术
BCR由两种蛋白质组成,重链和轻链,每一种蛋白质都由特定的基因片段编码而来。编码形成重链的基因片段位于第14号染色体上,我们知道每个B细胞都有两条14号染色体,一条来自于你的母亲,一条来自于你的父亲,而每个B细胞只能产生一种BCR或抗体,这就带来了一个问题:当有两组重链基因片段时,B细胞该如何选择让哪一组重链基因片段表达呢?
进化给出了一个绝妙的方案,两条染色体各自进行基因重排,首先选择一个可能的D和J片段,然后通过删除他们之间的DNA序列将他们两个片段相连接,然后再从众多V片段中选择其中一个V片段,通过删除V片段与DJ片段之间的DNA序列将V片段连接到DJ片段上。紧挨着J片段的是编码各种恒定区的基因片段。所选择的 V、D、J 基因片段和恒定区基因片段在14号染色体上彼此相邻。

接下来,必须对重排的基因进行检测。我们知道,当核糖体遇到三个终止密码子中的任何一个时,蛋白质翻译将停止,因此如果基因片段没有正确地连在一起,蛋白质翻译体系就遇到终止密码子,并终止蛋白质翻译过程。如果其中一条染色体发生了正确的重排,该染色体就可用来构建众链蛋白,成功表达的众链蛋白随后被转运到细胞表面,而结束的信号会传递给另一条染色体,使其停止重排。如果两条染色体都不能正确组装基因片段,不能表达受体的B细胞是完全没用的,因此它会选择自杀。
这样每个存活下来的成熟B细胞,都会产生而且只能产生一种特定的BCR,也就是由它最终选定的那一套V D J重链基因片段和那一套V J轻链基因片段编码的受体。但是,由于在制造每个B细胞的重链和轻链时,都采用了这种混搭策略,从成千上万种V、D、J片段中进行选择和组合,这就使得不同B细胞表面的受体呈现出极其惊人的多样性。
BCR信号转导
BCR识别了与它匹配的抗原后,要把这种识别信号传递到细胞核,但是BCR的重链胞内部分太短了以至于不能直接传递抗原刺激信号。这时候该怎么办呢?B细胞非常聪明,为使BCR能顺利传递它所看到的信号,B细胞配备了两种辅助蛋白,分别是Igα和 Igβ,它们与重链蛋白结合并伸入细胞内部。所以实际上,完整的B细胞受体包括两部分:细胞外能识别抗原但不能传递信号的重链轻链部分,以及能传递信号的Igα和Igβ蛋白。
为了产生足够强的激活信号,B细胞表面上许多BCR必须得紧密聚集到一起。当抗原与B细胞接触后,B细胞表面的多个BCR复合物将会与抗原结合而发生聚合,我们把这种现象称为“交联”。交联主要有几种情况:第一种,抗原本身有很多相同的表位,BCR就能同时抓住好几个表位,把自己拉到一起。第二种,很多抗原分子聚集在入侵者表面,比如细菌表面有很多相同的蛋白质分子。只要BCR能识别其中一种蛋白质的表位,就能把好几个BCR拉到一起。第三种,BCR结合到已经聚集在一起的抗原表位上,也能被拉到一起。
不管通过哪种方式,BCR的交联对于B细胞的激活来说,是绝对必需的!为什么呢?因为只有当足够多的Igα,Igβ分子聚集在一起时,它们才能有效地与细胞内的酶相互作用,启动一系列的酶联反应,就像多米诺骨牌一样,最终把BCR参与的信号传递到细胞核。所以,BCR的交联是B细胞激活的必要条件。
除B细胞受体及其Igα和Igβ外,B细胞表面还有另一种蛋白质也可能在信号传递中发挥重要的作用。这种蛋白质是一种补体受体,它能与正在修饰入侵者的补体片段相结合。因此,对被调理的抗原而言,B细胞上有两种能与其结合的B细胞受体,识别抗原上特异性表位的BCR和识别被修饰抗原的补体受体。被调理的抗原这时就像一个“钳夹”,把BCR和补体受体聚集在一起。当BCR和补体受体通过被调理的抗原以这种方式相交联时,BCR所要传递的信号被大大地放大了。补体受体对信号传递有重要的影响:补体受体参与后,激活B细胞所需的BCR数量可减少至少100倍。在入侵者进攻的起始阶段,可与B细胞受体交联的抗原数量有限时,这个体系的功能尤为重要。通过B细胞的共受体识别被调理的入侵者,有助于使B细胞对先天免疫系统已经确认为危险的抗原更加敏感。
B细胞激活
我们体内的大多数B细胞从来没有遇见过他们的同源抗原,这些B细胞被我们通俗地称为初始B细胞。初始B细胞可以通过两种方式被激活,一种完全依赖于辅助T细胞的帮助,称为T细胞依赖型激活,一种基本上不依赖T细胞的帮助,称为非T细胞依赖型激活。
T细胞依赖型激活需要两种信号。第一种信号由 BCR与相关的信号分子的聚集而提供,然而第一种信号并不足以激活B细胞,因此需要第二种信号,称为共刺激信号,这通常由辅助性T细胞提供。目前研究最清楚的共刺激信号是B细胞和辅助T细胞的直接接触。激活的辅助T细胞表面存在CD40L,B细胞表面存在CD40,当 CD40L 与B细胞表面的CD40 蛋白结合且BCR已经交联时,B细胞就被激活,从而开始传递刺激信号。
对于某些特定的抗原,初始B细胞可以在很少或没有T细胞的帮助下就能被激活,这种激活模式称为非T细胞依赖型激活。这些抗原通常具有重复的表位,可以交联大量的BCR。事实上,大量B细胞受体的聚集作用可以部分取代 CD40L 的共刺激作用。但是,要想完全激活B细胞并产生抗体,对于非T细胞依赖型激活方式而言,还需要一个明确的“危险信号”,这通常通过炎症环境或模式识别受体来提供。
非T细胞依赖性激活的优点是B细胞无需等待辅助细胞的激活,而可以直接加入战争,从而能更快地产生抗体应答。此外,辅助T细胞只识别由MHC2类分子提呈的多肽并被激活,如果B细胞的激活必须要依靠辅助T细胞,那么整个适应性免疫系统仅能对付蛋白质类外来入侵物质。因此,大自然设计出了非T细胞依赖性激活的方式,让B细胞能够在没有辅助T细胞的帮助下也能顺利被激活,从而增加了适应性免疫系统可以对付的抗原的范围,不仅包括蛋白质,还包括了糖类、脂类和核酸。
B细胞一旦被激活并通过增殖建立起了它们的数量群,它们就准备好了生命的下一个阶段,成熟。成熟可以粗略地分成三个阶段:第一个阶段是“类型转换”,在这个阶段B细胞能改变它所产生的抗体的类型。第二个阶段是“体细胞高突变”,在这个阶段编码BCR的重排基因能经历突变和选择来增加BCR对其同源抗原的亲和力,最后一个是“职业决定”,在此期间B细胞会决定是变成一个抗体工厂,也就是浆细胞,还是成为一个记忆B细胞。
结语
B细胞是高度专业化、多样化的免疫细胞,其核心特征是BCR,通过复杂的基因重排和选择机制产生。BCR识别抗原后,通过Igα和Igβ传递信号,并且需要交联和共刺激信号。 B细胞激活需要严格调控,确保安全性和有效性。它在成熟过程中经历类别转换、亲和成熟和命运抉择三个阶段。这一切都展示了适应性免疫系统惊人的复杂性和精妙设计。
参考书目:《How the immune system works》