引言
在过去的一个世纪里,已经开发出化疗和放疗等强效治疗方法来治疗癌症。不幸的是,这些方法通常缺乏足够的选择性,无法在避免不可耐受毒性的情况下使用足以根除癌细胞的高剂量。单克隆抗体能够提供显著扩大治疗窗口所需的高特异性水平,一些抗体甚至能够区分仅相差一个氨基酸或一个翻译后修饰的两个抗原。
几项关键技术的发展使得抗体能够用于治疗目的。首先,20世纪70年代开发的杂交瘤系统允许生产和筛选针对人类抗原的高特异性小鼠单克隆抗体。其次,将人抗体恒定区移植到小鼠抗体可变区的能力产生了具有更好治疗效果和更少不良反应的嵌合抗体。第三,20世纪90年代转基因小鼠模型和噬菌体展示系统的使用使得能够生成针对癌症靶点的全人源抗体。随着这些技术进步和临床需求的增加,过去二十五年见证了基于抗体的新疗法的爆炸式增长。自1997年以来,已有超过五十种用于肿瘤学应用的基于抗体的治疗剂获得美国食品和药物管理局(FDA)和欧洲药品管理局(EMA)的批准。除了基于抗体的治疗剂外,抗体片段还用于生成用于T细胞工程的嵌合抗原受体。
-02-
一、抗体生成
一系列技术被用于识别靶向肿瘤抗原的治疗性单克隆抗体或抗体片段。这些技术包括用靶抗原免疫啮齿动物、噬菌体或酵母展示等展示技术、从人和转基因小鼠中进行单B细胞克隆,以及从骆驼科动物中识别单域抗体。具有临床实用性的抗体的进一步开发涉及抗体工程,使其逐渐与人类免疫系统更加兼容。
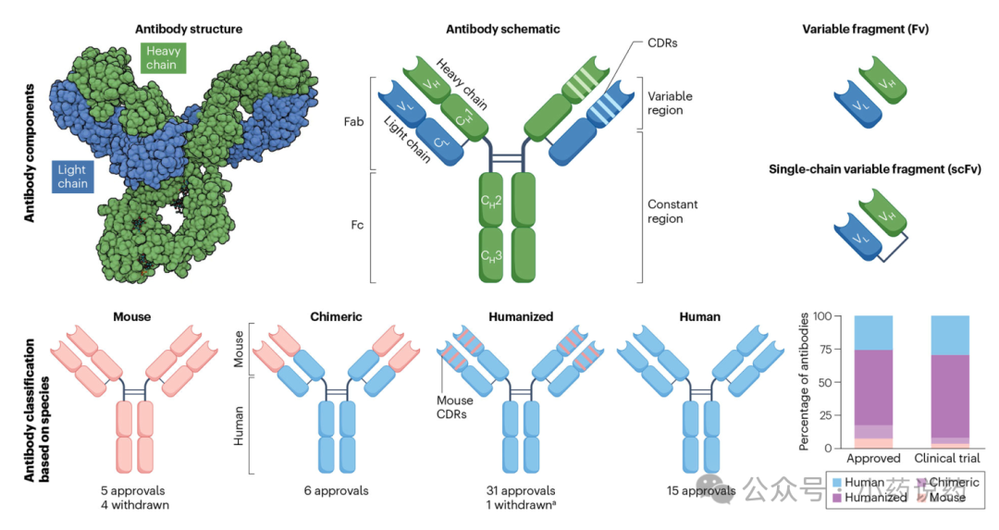
嵌合和人源化抗体
单克隆抗体传统上是通过使用1975年Cesar Milstein和Georges Kohler开发的杂交瘤技术生产的。在此过程中,用特定抗原免疫后,从小鼠脾脏或淋巴结中分离B细胞,然后与骨髓瘤细胞系(如Sp2/0)融合,形成称为杂交瘤细胞系的杂交细胞。通常通过酶联免疫吸附测定筛选蛋白质结合,通过流式细胞术筛选细胞结合来筛选杂交瘤克隆。杂交瘤技术稳健且经济高效地产生了一系列小鼠单克隆抗体。然而,注入人体的小鼠抗体会产生人抗鼠抗体免疫反应,限制了其疗效并增加了不良反应。因此,大多数获得监管批准的小鼠抗体后来都被撤市。Blinatumomab是一种使用两个小鼠来源的单链可变片段格式且缺乏Fc片段的双特异性T细胞衔接器,是唯一在临床得到广泛采用的鼠源抗体。
为了减少患者免疫反应的诱导,将小鼠抗体的恒定区替换为其人源对应部分以产生嵌合抗体。利妥昔单抗和西妥昔单抗是最早产生的嵌合抗体之一,通过将小鼠抗体2B8和225的可变区连接到人免疫球蛋白G1重链和人κ轻链恒定区而生成。然而,具有鼠源可变区的嵌合抗体仍可被识别为外来物,导致可能清除治疗性抗体的人抗嵌合抗体免疫反应。这可能会限制嵌合抗体的重复应用并阻碍其临床开发。为了进一步减少不必要的免疫反应,通过20世纪80年代开发的称为“人源化”的过程增加了小鼠单克隆抗体的人源含量。人源化涉及仅将小鼠抗体的互补决定区移植到人抗体的框架区。曲妥珠单抗是最早开发的人源化抗体之一,通过将小鼠HER2抗体mumAb4D5的CDR移植到人抗体框架中而开发,类似的策略被用于开发奥比妥珠单抗、帕博利珠单抗和阿特珠单抗。
除了小鼠杂交瘤技术外,还开发了兔杂交瘤用于生产针对肿瘤抗原(如间皮素)的兔单克隆抗体。CDR移植方法已被用于人源化兔单克隆抗体,例如针对间皮素的YP218,用于临床开发。监管机构批准的大多数治疗性抗体都是使用杂交瘤方法生成,然后工程化为嵌合或人源化形式,并广泛用于癌症治疗。
人源抗体
借助20世纪90年代开发的两项技术,即人抗体噬菌体展示和人抗体表达转基因小鼠模型,实现了生成全人源抗体的最后一步。为了生成产生人源抗体的转基因小鼠,将人Ig基因座或可变区插入小鼠基因组,同时破坏小鼠Ig基因。从用靶抗原免疫的小鼠中分离B细胞用于单B细胞克隆和测序。转基因小鼠平台XenoMouse、VelocImmune和HuMab产生了九种获批的靶向癌症的人源抗体:达雷妥尤单抗、伊匹木单抗、纳武利尤单抗、奥法木单抗、帕尼单抗、度伐利尤单抗、西米普利单抗、替索妥单抗和瑞拉利单抗。这些抗体成功的临床开发导致了多个转基因小鼠平台的迅速兴起,例如KyMouse、OmniRat、H2L2 Mouse、Trianni Mouse和RenMab。
展示技术为人源抗体开发提供了替代平台。使用噬菌体展示,可以从多样性超过10¹⁰的大型文库中分离人scFv或Fab片段。噬菌体展示技术通常使用M13丝状噬菌体来表达与噬菌体pIII外壳蛋白融合的抗体片段,如scFv。用固定在微孔板或珠子上的靶抗原筛选噬菌体文库。洗去非特异性噬菌体结合物,收获剩余的特异性结合物用于下一轮筛选。通常需要三到五轮筛选来分离特异性结合物。这个过程称为噬菌体淘选,它模拟了体外的免疫选择。使用下一代测序的新方法有助于快速识别稀有结合物。
Dyax的噬菌体展示系统产生了三种获得监管批准的全人源抗体:雷莫西单抗、奈西妥珠单抗和阿维鲁单抗。此外,由Dyax、Cambridge Antibody Technology、MorphoSys等公司产生的多种噬菌体展示来源的人源抗体目前正在进行临床试验。转基因小鼠和噬菌体展示技术都有其独特的优势。通常,转基因小鼠平台倾向于产生具有更理想生物物理特性和在临床试验中表现更好的抗体,而噬菌体展示允许选择靶向特定表位的抗体。除了噬菌体展示外,细胞表面展示、核糖体展示和mRNA展示技术也已用于抗体生成。酵母和哺乳动物细胞表面展示允许在真核细胞表面表达重组抗体片段。这使得能够通过流式细胞术分离抗原特异性细胞并发现高亲和力结合物。
比较嵌合、人源化和人源抗体
嵌合、人源化或全人源抗体生产的目标是减少针对抗体的免疫反应,这种反应带有重复给药导致中和和不良反应的风险。对各种抗体疗法进行的大型人类临床试验分析显示,使用嵌合、人源化和人源抗体时,免疫原性逐渐下降。然而,该研究未检验较低的免疫原性是否会导致更优的肿瘤消退或患者生存期。几种FDA和EMA批准的嵌合、人源化和人源抗体靶向常见的癌症抗原,如CD20、HER2和EGFR,尽管表位不同且结合特性不同。比较靶向CD20、HER2和EGFR的抗体的III期临床试验已证明相似的总生存期和相似的治疗相关停药率,表明治疗效果相当。然而,显著的HAMA免疫反应阻碍了靶向双唾液酸神经节苷脂GD2的小鼠抗体的开发,并且在自身免疫性疾病患者中使用利妥昔单抗观察到了强烈的HACA免疫反应。对HACA免疫反应的担忧,加上简化人源化和人源抗体生成的技术进步,增加了这两种格式的利用。进入当前临床试验的抗体要么是人源化产品,要么是全人源产品,并且根据之前的试验结果,人源化和人源抗体在患者中往往表现出相似的治疗效果。
-03-
二、抗体疗法的不同形式
根据其结构和功能机制,抗体疗法可分为三大主要格式:单特异性抗体、双特异性抗体以及与载荷偶联的抗体。
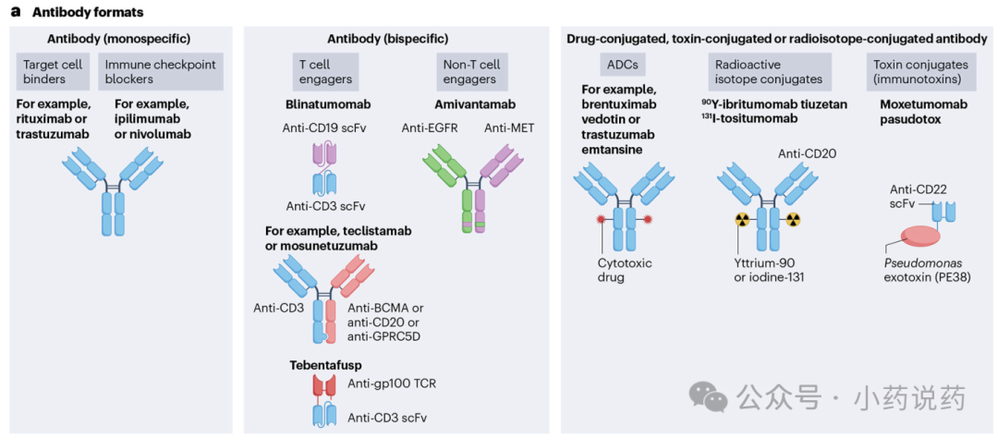
单特异性抗体格式
单特异性抗体格式涉及结合靶抗原的全长免疫球蛋白。在五种免疫球蛋白同种型中,只有IgG与新生儿Fc受体结合,导致半衰期较长。靶向癌症的抗体利用IgG同种型来利用这种延长的半衰期,并且通常每21天给药一次。大多数FDA和EMA批准的抗体以及开发中的抗体都使用单特异性IgG抗体格式。IgG抗体以四种亚类存在,大多数治疗性抗体使用IgG1亚类。单特异性抗体格式的靶抗原是细胞表面蛋白,主要是实体瘤中过表达的生长因子受体。对于血液系统恶性肿瘤,抗体通常靶向由不同免疫细胞亚群表达的细胞表面糖蛋白。
作用机制:抗体与癌细胞结合通过多种机制导致癌细胞死亡。直接阻断来自生长因子的生存信号和阻断血管生成导致肿瘤血供破坏是这些抗体导致实体瘤消退的两个主要机制。相比之下,血液系统恶性肿瘤中靶向的CD标记物通常在促进癌细胞存活方面没有实质性作用,相反,抗体通过募集和激活免疫效应细胞来诱导细胞毒性。
每个IgG抗体亚类赋予抗体不同的效应功能。IgG1亚类Fc段与巨噬细胞和自然杀伤细胞表达的激活Fcγ受体具有强亲和力。FcγR结合导致巨噬细胞和NK细胞介导的抗体依赖性细胞毒性和抗体依赖性细胞吞噬作用,直接杀死癌细胞。最早获得FDA批准的两种抗体是靶向恶性B细胞表达的CD20的利妥昔单抗和靶向乳腺癌细胞过表达的HER2的曲妥珠单抗,这两种抗体都使用常见的IgG1同种型。IgG1诱导ADCC和ADCP的能力最终导致癌细胞死亡,并且可以通过Fc工程进一步增强。IgG1亚类还结合补体蛋白C1q,通过补体依赖性细胞毒性导致癌细胞死亡。目前,市售的单克隆抗体主要由IgG1组成。除了效应功能外,IgG亚类的选择还基于结构稳定性、循环半衰期、制造和监管批准经验、缺乏免疫原性或意外副作用,以及公司开发组合中特定IgG亚类的可用性。
单特异性抗体Fc工程化:大多数IgG1抗体效应功能由Fc结构域介导。数十年的研究增加了我们对Fc结构域与不同Fc受体相互作用的理解,并帮助设计了具有理想效应功能的抗体。在临床前研究中显示活性增加的常见Fc结构域修饰包括去岩藻糖基化或关键残基的氨基酸取代,两者都导致增强的ADCC和ADCP。
免疫检查点抑制剂
在过去十年中,一类完全不同的靶向免疫细胞调节检查点的单特异性抗体显示出显著的临床疗效。目前有11种FDA和EMA批准的免疫检查点抑制抗体正用于治疗超过二十种不同类型的癌症,包括肺癌、黑色素瘤、肾细胞癌、头颈部鳞状细胞癌,预计未来还会有更多此类抑制抗体获得批准。免疫检查点抑制剂在大多数癌症类型中显示出20-30%的反应率。然而,某些恶性肿瘤,如霍奇金淋巴瘤和皮肤癌、微卫星不稳定性高癌症、PDL1表达升高的癌症、或高突变负荷的癌症,以及一些与病毒相关的癌症,反应率显著更高。
作用机制:免疫检查点阻断抗体抑制负向调节T细胞的通路,从而重振细胞毒性T细胞以杀死癌细胞。在临床试验中正在研究的超过20个免疫检查点中,治疗性抗体已获得FDA或EMA批准的三个蛋白质或通路是CTLA4、PD1-PDL1和LAG3。CTLA4、PD1和LAG3的表达在T细胞刺激后被诱导,其主要目的是限制T细胞活化的程度。
PD1阻断抗体是迄今为止使用最广泛的免疫检查点抑制剂。七种获批的PD1阻断抗体和两种目前正在进行临床试验的抗体使用IgG4格式,与IgG1同种型相比,该格式不能有效激活补体级联反应且Fc受体结合较弱。因此,IgG4格式可能保护表达PD1的效应T细胞免于被ADCC或CDC意外杀死。类似的IgG4格式也用于靶向LAG3的抗体瑞拉利单抗。所有IgG4抗体都携带S228P突变以防止Fab臂交换。
免疫检查点抑制剂毒性:免疫检查点阻断抗体通过一般免疫激活介导癌细胞杀伤,这有时可能会错误地针对健康组织。免疫检查点抑制抗体具有非常独特的不良反应谱,称为免疫相关不良事件。这些irAE包括各种表现,包括皮肤、胃肠道、肝脏、内分泌、肺部、神经和心脏事件。虽然不常见,但偶尔可能因免疫检查点抑制而产生严重且可能致命的毒性。在许多情况下,需要使用糖皮质激素、肿瘤坏死因子拮抗剂、霉酚酸酯或其他免疫调节剂进行临时免疫抑制来控制irAE。
双特异性抗体格式
第二种格式包括多样化的双特异性抗体类别。与单特异性抗体不同,双特异性抗体结合两种不同的抗原或表位。抗原可以位于同一靶细胞上或不同细胞上。靶向两种不同细胞的双特异性抗体大多是T细胞衔接器,将癌细胞与效应T细胞交联;因此,它们被称为T细胞衔接器。交联后,效应T细胞被激活,通过释放细胞毒性颗粒和淋巴因子杀死结合的靶癌细胞。另一类双特异性抗体参与同一靶细胞表达的两种不同抗原,例如两种不同的生长因子受体。此类双特异性抗体通过阻断靶生长因子受体的增殖信号以及激活针对癌细胞的NK细胞和巨噬细胞来杀死靶细胞。
双特异性T细胞衔接器:博纳吐单抗在2014年获得监管批准,导致了此类基于抗体的治疗剂的爆炸性增长。原型TCE双特异性博纳吐单抗是一种小的54 kDa融合蛋白,大小约为IgG抗体的三分之一。博纳吐单抗由双特异性抗体所需的最基本元件组成;一个靶向癌症的scFv通过甘氨酸-丝氨酸肽连接子与一个结合T细胞的scFv连接。博纳吐单抗通过抗CD19 scFv结合B细胞,同时通过抗CD3 scFv连接并激活T细胞,导致B细胞死亡。博纳吐单抗在一系列B细胞恶性肿瘤中显示出活性,并获得批准用于治疗B细胞前体急性淋巴细胞白血病。
博纳吐单抗的基本设计被后续获得监管批准的TCE双特异性抗体所采用。制药公司通常对使用独特架构和方法组装双特异性抗体的独特TCE格式进行商标注册。常见格式包括双特异性T细胞衔接器、Duobody、DART和Xmab,博纳吐单抗使用BiTE格式。Tebentafusp是一种独特的TCE双特异性格式,名为ImmTAC,它将靶向黑色素瘤细胞表达的gp100-HLA-A02复合物的亲和力增强的TCR与抗CD3 scFv连接起来。Tebentafusp在转移性葡萄膜黑色素瘤患者中显示出优越的总生存期,于2022年获得批准。在过去2年中,六种靶向血液系统恶性肿瘤的新双特异性抗体获得监管批准。这些包括teclistamab和elranatamab、talquetamab用于多发性骨髓瘤,以及mosunetuzumab、epcoritamab和glofitamab用于B细胞淋巴瘤。然而,TCE双特异性抗体在实体瘤中的批准滞后。然而,针对实体恶性肿瘤的TCE双特异性抗体临床试验激增,为这一策略将扩大其应用范围带来了希望。最近关于TCE双特异性抗体靶向DLL3在小细胞肺癌中的阳性I期试验报告令人鼓舞,表明实体瘤障碍将被打破。
靶向同一细胞上不同抗原的双特异性抗体:双特异性抗体也被设计为结合靶癌细胞上的两种不同抗原或表位,而不参与效应T细胞。它们的抗癌作用是通过阻断两个增殖信号通路介导的,从而最大化抗肿瘤活性。除了其受体阻断活性外,这些双特异性抗体还可以被工程化以包含功能性的IgG1 Fc结构域,使它们能够通过非T细胞基础的免疫效应通路(如ADCC、ADCP和CDC)杀死癌细胞。阿米万他单抗是第一个靶向癌细胞表达的EGFR和MET的受体阻断双特异性抗体,并获得监管批准用于治疗具有外显子20插入突变的非小细胞肺癌。阿米万他单抗比单受体结合抗体的组合更有效地阻断驱动肺癌亚群的EGFR和MET通路信号传导。此外,阿米万他单抗的IgG1 Fc结构域被工程化为具有低岩藻糖水平,这增强了FcγRIIIa结合和NK细胞介导的ADCC。类似的双特异性设计被扎尼达他单抗采用,该抗体用每个Fab臂结合两个不同的HER2表位。这些臂靶向HER2亚结构域2和亚结构域4,分别是帕妥珠单抗和曲妥珠单抗的结合位点,这两种靶向HER2的抗体被批准用于治疗HER2+乳腺癌。临床前研究表明,与曲妥珠单抗、帕妥珠单抗或曲妥珠单抗和帕妥珠单抗的组合相比,扎尼达他单抗增强了HER2聚集、受体下调并增加了CDC介导的HER2+癌细胞杀伤。在一项I期试验中,扎尼达他单抗在一系列HER2+实体瘤中显示出鼓舞人心的活性。
双特异性抗体毒性:TCE双特异性抗体具有相似的不良反应,因为它们使用通过T细胞活化杀死靶细胞的共同抗癌机制。毒性水平与携带TCE双特异性抗体靶点的细胞的总身体负担有些相关。TCE双特异性抗体诱导以发热、不同程度的缺氧、低血压和偶尔多器官衰竭为特征的全身性炎症,统称为细胞因子释放综合征。神经毒性是与TCE双特异性抗体相关的另一种独特不良反应,表现为意识模糊和震颤,以及言语和行为的改变。CRS和神经毒性的病理生理学不完全清楚,可能与细胞因子水平升高有关。接受TCE双特异性抗体治疗的患者需要密切监测,并可能需要使用糖皮质激素或抗细胞因子药物(如IL-6中和抗体托珠单抗)进行免疫抑制。此外,已经实施了几种策略来预防或减少CRS和神经毒性,包括逐渐上调TCE双特异性抗体剂量、预防性使用糖皮质激素,以及在开始TCE双特异性抗体治疗前使用化疗减少肿瘤负荷。受体阻断双特异性抗体不能激活T细胞,因此不会诱导通常在TCE双特异性抗体中观察到的CRS或神经毒性。受体阻断双特异性抗体与单特异性抗体对应物的不良反应相似,例如阿米万他单抗引起的皮疹以及扎尼达他单抗引起的心力衰竭。
偶联抗体形式
第三种主要格式涉及与毒性载荷连接的抗体,例如细胞毒性药物、细菌或植物毒素,或放射性同位素,这增强了抗体杀死癌细胞的能力。在该组中,ADC是迄今为止使用最广泛的格式,而毒素偶联和放射性同位素偶联抗体尚未被广泛采用。
抗体药物偶联物:ADC是通过将肿瘤靶向抗体与细胞毒性药物连接而构建的。ADC分子与细胞表面抗原的结合导致其内化,随后在细胞内释放细胞毒性药物。这使得细胞毒性药物能够选择性地递送至癌细胞,同时保留大部分健康组织。ADC的关键组成部分包括肿瘤靶向抗体、细胞毒性药物和连接抗体与细胞毒性药物的连接子。ADC的成功取决于这些关键组成部分的最佳选择,以及用于将连接子连接到抗体的偶联方法,这通常决定了药物抗体比。
大多数ADC使用人源化或人源IgG1作为肿瘤靶向抗体。如上所述,使用IgG1的流行是由于其约21天的长血浆半衰期,以及其结合Fc受体导致通过ADCC和ADCP增强靶细胞杀伤的能力。两种ADC,吉妥珠单抗和奥英妥珠单抗,使用IgG4,其对FcγRII和FcγRIII的亲和力较低,从而限制了ADCP,同时可能由于通过Fc受体非特异性摄取ADC进入免疫细胞减少而降低毒性。
大多数连接子将细胞毒性药物连接到IgG1抗体骨架上的随机赖氨酸或半胱氨酸残基。有效的连接子最大限度地减少细胞毒性药物在血流中的早期释放,同时促进活性药物在优选靶向位置的受控释放。连接子大致分为可裂解和不可裂解。多数ADC中使用可裂解连接子,例如肽连接子、腙连接子、二硫键连接子或CL2A连接子。为药物连接开发的第一个连接子之一是使用腙键的可裂解连接子。该连接子用于将抗肿瘤抗生素卡奇霉素连接到ADC吉妥珠单抗和奥英妥珠单抗。腙键被设计为在靶细胞溶酶体内的酸性条件下分解以释放卡奇霉素。然而,腙键可以在血浆中发生水解,导致药物分子意外释放,引起全身毒性。第二种可裂解连接子是肽连接子,需要溶酶体内组织蛋白酶B介导的选择性裂解。四种ADC使用mc-VC-PABC二肽连接子,该连接子由Seagen首次开发用于生成ADC Brentuximab,该ADC使用与微管抑制剂单甲基澳瑞他汀E连接的靶向CD30的抗体。Brentuximab联合化疗在几种不同淋巴瘤亚型的患者中显示出显著疗效,被认为是霍奇金淋巴瘤和CD30+外周T细胞淋巴瘤的标准一线治疗。Brentuximab的成功导致了mc-VC-PABC二肽连接子的广泛采用。Seagen与罗氏、安斯泰来和Genmab合作,使用mc-VC-PABC二肽连接子分别开发靶向CD79B、nectin-4和TF的ADC。第三种可裂解连接子是二硫键连接子,其中连接子裂解由细胞内高浓度的谷胱甘肽介导。具有二硫键连接子的ADC包括mirvetuximab,该药在卵巢癌患者的III期试验中显示出生存获益。与可裂解连接子相比,不可裂解连接子对各种分解机制具有抵抗力。载荷在靶细胞溶酶体内抗体降解后释放。两种ADC,曲妥珠单抗美坦新和belantamab mafodotin,使用不可裂解连接子进行药物连接。临床前研究表明,由于不可裂解连接子的血浆稳定性增加,对非靶细胞的毒性较低。然而,可裂解和不可裂解连接子尚未在临床试验中进行直接比较,一种相对于另一种的优势仍不清楚。
肿瘤细胞杀伤过程由连接到肿瘤靶向抗体的细胞毒性药物执行。该药物通常是一种具有高细胞杀伤效力的小分子,通过三种机制之一诱导细胞死亡:直接DNA损伤、破坏微管网络或抑制拓扑异构酶活性。目前使用的药物具有广泛不同的IC₅₀,从5 nM到5 pM。然而,ADC的效力取决于药物的IC₅₀和DAR。因此,具有高效力PBD载荷和相对较低DAR的loncastuximab tesirine与具有较低效力载荷Dxd但相对较高DAR的曲妥珠单抗德鲁替康都能够在几种体内模型中诱导完全肿瘤消退。具有高DAR和新型连接子设计的ADC的益处也在临床试验中观察到。曲妥珠单抗美坦新和曲妥珠单抗德鲁替康共享相同的靶向HER2抗体。然而,曲妥珠单抗美坦新携带通过不可裂解连接子连接的微管抑制剂DM1,而曲妥珠单抗德鲁替康利用通过可裂解连接子连接的药物Dxd。在比较两种ADC的III期试验中,曲妥珠单抗德鲁替康显示出更高的反应率和总生存期。此外,曲妥珠单抗德鲁替康延长了表达低水平HER2的乳腺癌患者的生存期,使其成为第一个对HER2低乳腺癌有效的ADC。最近,fam-trastuzumab deruxtecan在一系列具有高HER2表达的实体瘤中显示出肿瘤消退,并获得了首个FDA批准的肿瘤不可知HER2导向疗法。
ADC毒性:ADC被认为是靶向药物,比传统细胞毒性化疗耐受性更好。然而,大多数患者在使用ADC时会出现某种形式的毒性。大多数ADC共有的毒性包括输注反应、血细胞减少、感染、肝酶升高、胃肠道症状和胚胎-胎儿毒性。某些毒性与特定载荷的使用有关,例如微管抑制剂引起的周围神经病变和卡奇霉素引起的肝毒性。其他毒性在靶向共同抗原的ADC中普遍存在。例如,靶向HER2的ADC可引起心力衰竭并介导间质性肺病。
抗体-毒素偶联物或免疫毒素:免疫毒素有两个组成部分,一个靶向抗体或抗体的Fv,以及一个通常来自细菌或植物毒素的细胞毒性蛋白质。靶向抗体或Fv结合靶细胞,允许选择性递送毒素。毒素来自细菌,如假单胞菌外毒素A和白喉毒素,或来自植物,如蓖麻毒素。理论上,任何用于生成ADC的靶向抗体都可用于免疫毒素。然而,尽管过去几年有12种ADC获批,但只有一种免疫毒素莫昔妥莫单抗帕苏多托获得FDA和EMA批准用于治疗毛细胞白血病。
莫昔妥莫单抗是一种融合蛋白,将CD22结合的Fv与截短的假单胞菌外毒素A组合,由美国国家癌症研究所的I. Pastan开发。进入靶细胞后,PE38毒素与延伸因子2结合并阻断蛋白质合成。莫昔妥莫单抗在毛细胞白血病患者的临床试验中显示出41%的高完全缓解率,并于2018年获得监管批准。然而,由于临床使用率低,莫昔妥莫单抗的生产于2023年停止。
莫昔妥莫单抗和其他研究中的免疫毒素面临若干挑战,包括导致治疗效果丧失的高免疫原性,以及导致毛细血管渗漏综合征和溶血性尿毒症综合征等毒性的狭窄治疗窗。莫昔妥莫单抗采用率低也可能是因为给药的复杂性,包括毒性预防、治疗前和治疗后水化、需要安全监测以避免不良反应以及毛细胞白血病替代疗法的可用性。尽管过去有几项临床试验评估了靶向BCMA、间皮素和其他癌症相关抗原的免疫毒素的疗效,但目前没有一种处于监管审查阶段,也没有预计在不久的将来获得批准。
抗体-放射性同位素偶联物:抗体-放射性同位素偶联物由与放射性同位素连接的靶向抗体组成。放射性同位素发射α粒子或β粒子,导致靶细胞中的DNA链断裂,从而导致细胞死亡。抗体-放射性同位素偶联物不需要内化即可诱导细胞死亡,这提供了相对于ADC和免疫毒素的独特优势,包括旁观者效应。α粒子是大的带正电粒子,由两个质子和两个中子组成,有效范围相对较短。相比之下,β粒子是小的带负电电子,有效范围更长,但DNA损伤能量较低。β粒子发射体的例子包括碘-131、镥-177和钇-90。两种FDA和/或EMA批准的抗体-放射性同位素偶联物,⁹⁰Y-替伊莫单抗和¹³¹I-托西莫单抗,将β粒子发射放射性核素连接到靶向CD20的抗体上,用于治疗B细胞淋巴瘤。⁹⁰Y-替伊莫单抗和¹³¹I-托西莫单抗在临床试验中显示出65-80%的总体反应率和20-30%的完全反应率。两项研究都观察到一些可预测的治疗相关不良反应,包括健康骨髓暴露于辐射导致的长期和严重血细胞减少、¹³¹I-托西莫单抗中放射性碘导致的甲状腺功能减退,以及继发性恶性肿瘤,如骨髓增生异常综合征和白血病。尽管反应率高,但这些药物并未实现广泛的临床应用,2012年只有75名患者接受了¹³¹I-托西莫单抗。结果,¹³¹I-托西莫单抗于2013年自愿撤市。⁹⁰Y-替伊莫单抗仍可用于复发或难治性低级别B细胞淋巴瘤患者,但报告表明只有少数患者接受这种治疗。批准数量有限和临床采用率低可能是因为需要多学科团队来开发和在临床部署这些分子。很少有中心具备这种能力,因此,抗体-放射性同位素偶联物的使用落后于ADC。
-04-
三、挑战与未来展望
肿瘤学药物开发是一个艰巨的过程,成功率为3-7%。缺乏针对肿瘤学基于抗体的治疗剂成功率的具体数据,但进入I期试验的治疗性抗体中约有18%会进入药物上市阶段。这些低数字反映了药物开发过程中遇到的许多障碍。治疗性抗体开发是一项资源密集的多步骤操作。它需要生成针对靶抗原的多种抗体,然后选择少数具有理想结合和生物物理特性的先导候选物。然后使用小鼠模型在体内测试先导抗体的疗效,使用非人灵长类动物测试毒性和药代动力学,然后进行首次人体临床试验。多种在小鼠模型中显示出有前景疗效的抗体由于在非人灵长类动物或首次人体试验中的毒性而未能进入临床开发。尽管存在这些障碍,但一些抗体正在显示出鼓舞人心的临床前和临床数据,并可能在未来几年改变患者护理。
新靶点:实体瘤占新发癌症患者的大多数。很大比例的实体瘤患者出现需要全身治疗的转移性疾病,而这很少能治愈。因此,转移性肺癌、结肠癌、胰腺癌、乳腺癌、前列腺癌、肝癌和胆管癌共同导致了大多数癌症死亡。然而,与血液系统恶性肿瘤相比,针对实体瘤的抗体开发滞后,主要是由于缺乏可靶向的抗原。不同的淋巴瘤亚型约占所有癌症死亡的3%,但有五个肿瘤抗原被治疗性抗体靶向。相比之下,导致约21%癌症死亡的肺癌只有一个可靶向的肿瘤抗原。此外,胰腺癌、前列腺癌和脑癌完全缺乏FDA和/或EMA批准的治疗性抗体。这种差异主要是因为淋巴瘤和骨髓瘤起源于正常的B细胞,后者表达独特的可靶向抗原。此外,根除正常B细胞不会导致不可耐受的后果。不幸的是,靶抗原发现在实体瘤中产生的线索很少。因此,新靶点的识别可能需要涉及对大量患者样本进行单细胞测序,以寻找在癌症和正常组织中差异表达的抗原。此外,最近的临床试验显示,靶向claudins、HER3、FGFR2B、DLL3、CTGF和STEAP1的抗体显示出令人鼓舞的结果。
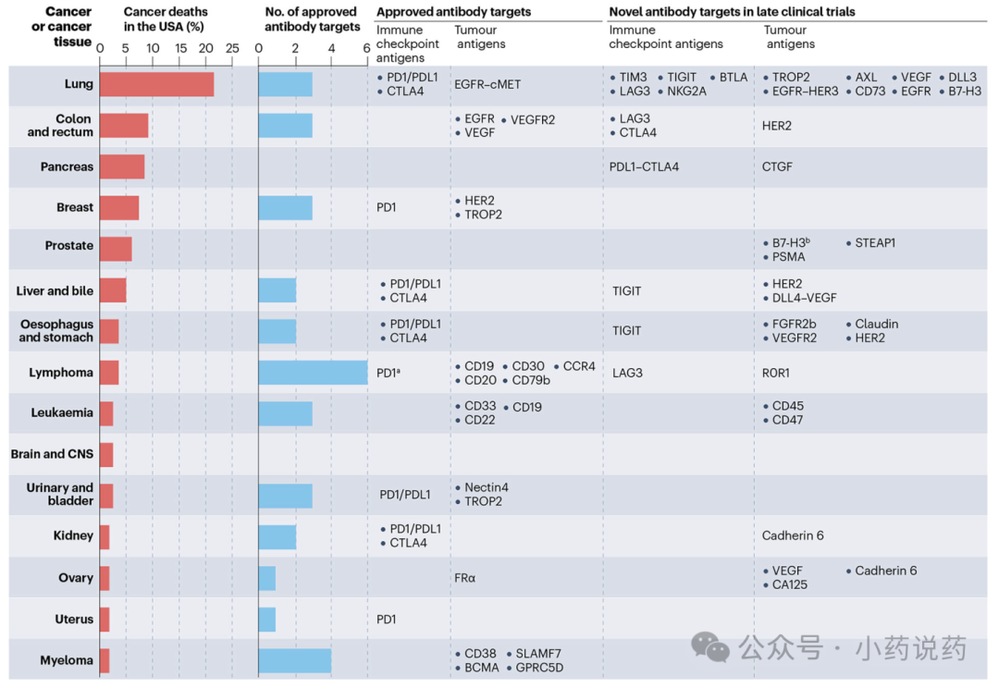
最后,规避缺乏肿瘤特异性抗原的一种创新方法是组合靶向多种抗原。大多数癌症缺乏在健康组织中不存在的真正特异性抗原的表达。因此,靶向仅在肿瘤中共表达而在健康组织中不共表达的抗原组合可能提供可行的治疗途径。利用需要两个或三个靶点激活细胞毒性机制的布尔逻辑AND门的临床前研究已证明肿瘤消退,但它们尚未在临床试验中进行测试。
一些细胞表面蛋白聚糖,如硫酸软骨素和硫酸乙酰肝素,是癌症和其他人类疾病中信号通路的重要调节剂。硫酸软骨素蛋白聚糖已被证明可促进肿瘤血管化并调节与肿瘤生长相关的信号转导通路,并且在多种癌症类型中表达。Glypicans是糖基磷脂酰肌醇锚定的细胞表面HSPG。几种glypicans如GPC1、GPC2和GPC3在癌症中过表达,表明它们是潜在的肿瘤相关抗原。针对GPC1、GPC2和GPC3的抗体和CAR T细胞已分别开发用于治疗胰腺癌、神经母细胞瘤和肝癌。
肿瘤特异性抗原:目前大多数获批的基于抗体的治疗剂靶向肿瘤相关抗原或组织特异性抗原。肿瘤相关抗原,如HER2,由癌细胞过表达,但也由一部分正常组织表达,尽管水平较低。癌症和正常组织之间表达水平的差异提供了治疗窗口。组织特异性抗原,如CD20,在癌细胞和正常细胞中以相似的水平表达。靶向此类组织特异性抗原的疗法旨在消除正常细胞和癌细胞,因为正常B细胞的耗竭在临床上是可耐受的。靶向CD19、CD20、CD38和BCMA的B细胞谱系癌症疗法代表了最成功的组织特异性治疗性抗体实例。
肿瘤特异性抗原的主要例子是新抗原,这些是由MHC分子呈递的突变肽。突变肽是携带氨基酸变化的蛋白质的短蛋白水解产物,这些变化源于突变,这是癌细胞的标志。来自突变癌症驱动基因(如BRAF、RAS、PIK3CA、TP53和异柠檬酸脱氢酶)的公共新抗原提供了对癌细胞极为独特的靶点,可用于造福患者。这些公共新抗原目前正被双特异性抗体靶向,并在临床前研究中显示出针对携带TP53和RAS突变的癌症的令人鼓舞的活性。
细胞内抗原:抗体无法穿透细胞膜,因此只能靶向细胞表面抗原。细胞内抗原可以与MHC分子结合呈递在细胞表面,使其可靶向。通过这种方式,tebentafusp靶向细胞内抗原gp100,该抗原在黑色素瘤中过表达,并与HLA-A*02:01结合呈递在细胞表面。现在正在开发抗体以靶向类似癌症特异性过表达的细胞内蛋白,如hTERT、MUC1、NY-ESO1、TARP、p53、WT1和PRAME,其中靶向PRAME在临床试验中显示出令人鼓舞的初步结果。
肿瘤微环境中的抗原:癌症在复杂的免疫细胞、成纤维细胞和细胞外基质混合物中发展,这被称为肿瘤微环境。目前的研究正在检查肿瘤微环境中免疫细胞的作用,如肿瘤相关巨噬细胞、树突状细胞和髓源性抑制细胞,这些细胞已知会驱动对免疫检查点抑制剂的耐药性。因此,耗竭或阻断这些免疫抑制细胞的功能可能使肿瘤对免疫检查点抑制剂敏感。
早期临床试验证明了通过靶向CSF1R、TREM1和TREM2进行抗体介导的TAM抑制的安全性和可行性。临床前研究还表明,使用激动性CD40单克隆抗体增强肿瘤微环境中树突状细胞的功能可以增强T细胞介导的肿瘤消退。所有对抗肿瘤微环境内免疫抑制状态的抗体目前正在II/III期试验中进行测试,这将决定它们的最终效用。
靶向低密度抗原:目前批准的ADC和双特异性抗体靶向相对丰富的抗原,如CD19、CD20和BCMA,每个癌细胞约有1,000至50,000个拷贝。靶向新抗原的主要挑战源于它们在癌细胞表面的低丰度,一些细胞表达1-10个靶抗原拷贝。除了新抗原外,有效靶向肿瘤相关或组织特异性抗原也可能受到其可变细胞表面密度的影响。此外,治疗压力也可以选择具有低靶抗原表达的癌细胞克隆。临床前研究表明,双特异性抗体能够杀死每个细胞表达低至1-10个抗原的癌细胞,而ADC需要1,000个或以上的拷贝才能杀死细胞。未修饰的全长抗体可能需要更高的靶抗原表达。靶向此类低密度抗原仍然是一个挑战,优化的抗体工程正在使其在临床前研究中有效靶向个位数水平的新抗原成为可能。
新型免疫检查点抑制剂:伊匹木单抗在2011年的批准迎来了免疫治疗时代。免疫检查点阻断剂具有前所未有的能力,可以在一些复发性实体瘤患者中诱导长期缓解,这导致了一系列临床试验的激增。FDA和EMA已批准11种不同版本的靶向CTLA4、PD1和PDL1的抗体,用于超过20种癌症类型的超过65种不同适应症。在CTLA4、PD1和PDL1靶向取得初步成功后,一系列额外的免疫检查点被探索,以诱导一系列癌症类型的缓解。
除了在黑色素瘤中靶向LAG3的瑞拉利单抗外,其他所有药物尚未在III期试验中证明获得监管批准所需的治疗活性。类似地,阻断CD47的巨噬细胞检查点抑制剂magrolimab在早期试验中显示出令人鼓舞的活性,但由于缺乏疗效,在MDS和AML患者中的III期试验被终止。目前新型免疫检查点阻断剂的表现低于预期,导致制药行业将重点放在PD1-PDL1领域。
新型抗体格式:所有全长治疗性抗体都使用IgG同种型。临床前研究和早期试验正在探索五聚体IgM格式,与IgG格式中的两个结合位点相比,它具有十个抗原结合位点。与靶向相同表位的IgG抗体相比,这提供了更高的结合亲合力。一个例子是IGM-8444,一种高亲合力五聚体IgM激动抗体,靶向死亡受体5,该抗体已在小鼠模型中显示出肿瘤消退,并已进入I期试验。
新型双特异性格式:TCE双特异性格式通过提高疗效同时减少不良反应而不断发展。目前有超过200种双特异性抗体处于临床开发阶段,其中大多数使用TCE方法靶向实体瘤。Glofitamab和xaluritamig在临床试验中均显示出疗效,并共享独特的2:1 T细胞衔接双特异性格式,具有对靶点的二价结合和对CD3的单价结合。与传统的1:1靶点与CD3结合设计相比,这种格式可能提供更高的肿瘤细胞杀伤能力。IgM格式TCE双特异性抗体由于使用五聚体IgM而具有更令人印象深刻的10:1靶点与CD3结合比。高靶点与CD3结合比设计允许高亲合力靶点结合,可能产生更高的靶点细胞毒性,同时减少细胞因子分泌。Imvotamab的I期试验已证明在晚期B细胞恶性肿瘤患者中具有几个完全缓解。
双特异性免疫检查点阻断剂:双特异性格式也被用于同时抑制两个免疫检查点,以增强T细胞活化,超越两种单特异性免疫检查点阻断剂组合所能达到的效果。靶向PD1xLAG3、PD1xCTLA4和PDL1xCTLA4的双特异性免疫检查点阻断剂在早期临床试验中显示出令人鼓舞的疗效。虽然最初的双特异性免疫检查点阻断剂组专注于抑制临床已确立的免疫检查点,但临床前研究正在调查类似的双特异性免疫检查点阻断剂是否可以通过靶向新型免疫受体来诱导抗肿瘤免疫。
三特异性抗体:三特异性抗体具有同时与三种不同抗原相互作用的能力。这种功能使三特异性抗体能够与两种不同的癌症相关抗原相互作用,同时在B细胞恶性肿瘤的背景下使用第三个结合域与细胞毒性T细胞结合。这种创新方法有助于减轻由于单一抗原丢失而引起的治疗耐药性,因为癌细胞必须同时下调两种抗原以逃避杀伤。或者,三特异性抗体被设计为与两个T细胞受体相互作用,从而向癌细胞传递更有效的激活信号。
纳米抗体:可能导致抗体工程重要范式变化的一个研究领域是单域抗体的开发。纳米抗体主要来源于骆驼科重链抗体,体积小,易于生产,稳定,并且能够穿透肿瘤或病毒抗原中的隐藏位点。纳米抗体通常从骆驼科来源的噬菌体展示文库中分离,最近也从转基因小鼠中分离。纳米抗体具有长的CDR3环,可以穿透抗原表面的空腔。第一个基于纳米抗体的靶向BCMA的CAR T细胞已获得FDA批准,并且与基于抗体的CAR T细胞相比显示出更高的疗效。靶向CD19或CD20和HER2的纳米抗体来源药物目前正在淋巴瘤和乳腺癌患者的临床试验中进行测试。纳米抗体是否会提供比当前基于抗体的药物更高的疗效仍有待观察。
可激活抗体:肿瘤微环境通常表现出较低的pH,含有活性蛋白酶酶,并具有正常组织缺乏的特定抗原组合。这些独特的特性被用来选择性地靶向癌细胞,使用可激活抗体,这些抗体根据环境条件的变化获得或失去功能。在抗体序列的某些位置添加组氨酸残基使其能够与靶抗原进行pH依赖性结合。具有这种掩蔽机制的抗体通常被称为前体药物疗法,正被用于改善PDL1、CTLA4、EGFR、CD166和CD71在一系列实体瘤和血液系统恶性肿瘤中的靶向。
新型偶联抗体:调节连接子设计和增加DAR可能会增强治疗益处。例如,具有高DAR的靶向细胞表面糖蛋白TROP2的ADC是第一个改善三阴性乳腺癌患者生存期的ADC。第一三共开发的创新四肽连接子用于曲妥珠单抗德鲁替康,掩盖了载荷DXd的疏水性,并允许均匀连接大量药物,同时保持良好的药代动力学并最大限度地减少药物在血浆中的过早释放。这种连接子-载荷组合现在正被几种接近批准或处于临床试验阶段的ADC采用。最后,新型药物载荷,如蛋白质降解剂、RNA聚合酶抑制剂、BCL-xL抑制剂、Toll样受体激动剂和干扰素基因刺激剂激动剂,预计将通过使用新型杀伤机制来改进ADC。
免疫毒素偶联物:免疫毒素的一个明显限制是抗毒素免疫反应的产生,这会缩短半衰期并降低疗效。目前的工作重点是去除被宿主免疫细胞识别的表位,以降低假单胞菌外毒素的免疫原性。这种修饰的免疫毒素可能会提高疗效,并允许免疫毒素靶向一系列恶性肿瘤。白介素-毒素偶联物的生成是该领域的另一项发展,并利用了癌细胞中白介素受体的过表达。Tagraxofusp是一种靶向IL3RA的细胞毒素,由IL-3与白喉毒素融合而成,在母细胞性浆细胞样树突状细胞肿瘤患者中显示出有效性。类似地,denileukin diftitox是一种IL-2R靶向细胞毒素,由IL-2与白喉毒素融合而成,在皮肤T细胞淋巴瘤患者中有效。然而,这些IL-毒素偶联物不被视为基于抗体的药物,因为它们使用受体配体而非scFv进行肿瘤靶向。
放射性同位素偶联物:在基于小分子和肽的放射性同位素偶联物在实体瘤中取得成功后,人们对放射性同位素偶联物重新产生了一些兴趣。将镥-177与结合前列腺特异性膜抗原的小分子或与结合生长抑素受体的肽类似物偶联,分别在前列腺癌和神经内分泌肿瘤患者中显示出显著的肿瘤消退,从而获得FDA批准。此类肿瘤靶向小分子-放射性同位素或肽-放射性同位素偶联物的行为类似于抗体-放射性同位素偶联物,但具有不同的结构和药代动力学。
-05-
结语
总之,基于抗体的治疗剂因其抗体与靶点结合所能提供的精确水平而在肿瘤学领域具有巨大前景。主要挑战仍然存在,但新靶点和各种新颖的设计和工程策略有望为许多这些挑战提供解决方案。获得监管批准的更有效的基于抗体的治疗剂的爆炸式增长可能会延续过去二十五年显示的趋势。
参考文献:
Cancer therapy with antibodies. Nat Rev Cancer. 2024 Jun;24(6):399-426.
公众号已建立“小药说药专业交流群”微信行业交流群以及读者交流群,扫描下方小编二维码加入,入行业群请主动告知姓名、工作单位和职务。
